Kleiber Rules!
Have you ever wondered why clinicians (and even some physiologists) relate
a number of measurements to body surface area? For example, they talk about
cardiac index (cardiac output divided by, yes, body surface area),
and urinary protein losses in nephrotic syndrome as 3.5g/1.73m2.
Why relate these measurements to area? Does this make sense?
Such an approach did make sense (at least on 23 July 1839) to the
mathematician Sarrus and the (heh) doctor of medicine Rameaux, who proposed
this "surface law" to the French Royal Academy. Theirs was a first primitive
attempt at scaling - trying to derive "laws" that apply whatever
the size of the organism. Clinicians, who usually have to deal with variations
in size over about two orders of magnitude (say 1 kilogram to about 100kg), have
been quite happy to accept the surface law and in so doing lag a hundred
years behind biologists, who cannot make do with sloppy approximation,
dealing as they do with organisms ranging in size from
a mycoplasma (so small that at physiological pH the mycoplasma contains
perhaps two hydrogen ions), to a hundred tons of blue whale. Biologists
consider weights ranging over twenty one orders of magnitude! It seems
inconceivable that there are scaling laws that apply across this vast
spectrum of life. But there are such "laws", and they have far reaching
implications for such diverse areas as ageing, athletic performance, and
weight-adjusted dosing.
What is the basis of Sarrus and Rameaux's argument? At first glance,
it seems quite sensible to argue that:
- As we increase the linear dimensions (L) of an organism, so its
surface area increases according to L squared, and weight according to L
cubed (Think of a sphere of radius r, with area 4pi.r2 and
volume 4/3pi.r3 and the result of replacing r with 2r). In
other words, weight increases more per unit increase in length than
does surface area.
- In any organism, energy loss must balance energy gain, so
(unless it is to freeze or burst into flame) heat loss (which should be
proportional to surface area) must be counterbalanced by appropriate
energy production (metabolism).
- Metabolism will then vary with surface area, and not weight.
QED? Not quite. Nature is often more subtle than nineteenth century
mathematicians. It is quite clear that small animals do live fast, short,
metabolically hectic existences compared to say elephants, but unfortunately
for Sarrus and Rameaux the rule isn't what they predict! We are about
to dip into the complex world of scaling.
Scaling Basics
The boundary where biology intersects mathematics is littered with the
intellectual corpses of frustrated clinicians who have tried unsuccessfully
to "crack into" the maths. A physician, I will not try to preach any
mathematical gospel and will stick to simple explanations. (Mirth-filled
mathematicians may suppress their laughter long enough to point out where
I've erred). We do however need some basic 'scalingspeak':
- isometric refers to the geometric similarity of bodies -
two bodies with the same proportions are isometric.
- The word allometric refers to bodies that are not
isometric, but that change with size according to a particular rule.
The term used is generally "allometric scaling", a useful internet
search term! We explore in detail rules for allometric scaling.
- dimensionless quantities are ratios between measurements
that (as their name suggests) don't have dimensions. For example,
if the height of something is 1 metre and its width is 2 metres,
we could make up a dimensionless quantity (let's call it the H:W ratio)
with a value of 0.5 that relates the two. We divide a length by a
length, so the result has no dimensions. Similarly, we might relate
the inertial and viscous forces in a fluid, and call this "Reynold's
number".
- dimensional correctness. Don't trust all the equations
you read in books. First, check that they are dimensionally correct.
An example:
Harris and Benedict derived equations predicting heat production
in men and women as follows:
- For men: Q = 66.4730 + 13.7516(w) + 5.0033(h) - 6.75505(y)
- Women:
Q = 655.0955 + 9.5634(w) + 1.8496(h) - 4.6756(y)
Where Q is heat production in kcal/day, w = weight in kilograms,
h = height in centimetres and y = age in years.
Consider these for a moment (quite apart from the improbable five
to seven digit accuracy). The equations might well give an
accurate prediction of energy production, but are they dimensionally
correct? Of course not. You can't add weight (dimension L3),
to height (dimension L) and obtain a sensible quantity. Kleiber
macerated these equations in 1932 (also rather amusingly pointing out that
by the above equations, a weightless woman has ten times the metabolic
rate of a weightless man)! Do we still use them?
- symmorphosis is a fancy word for a simple concept.
The concept, formulated by Taylor & Weibel in 1981, is:
Think about it - your grandfather's old truck from the 1940's
can survive a tornado, but your flashy new car dents
if you look at it. Motor manufacturers in the '40s overdesigned
like crazy, but now a new species of manufacturer has evolved
that knows just what it can get away with. Similarly, an organism
is unlikely to put energy into making overgrown bones that can resist
being stepped on by an elephant unless part of its daily life involves
being stepped on by elephants, an unlikely scenario! If your
cardiovascular system is never likely to be taxed more than say
ten times the demands on it at rest, not only will the system be
structured to cope with a maximum load of 10 times baseline, but
each and every component in the system will have this
same limitation! Likewise, it would be pointless for the lungs
to be able to provide say twenty times the resting oxygen usage
if the cardiovascular system could only distribute ten times
that basal amount.
- scale independent properties refer to things
that don't change with size. Muscles generate the same
power and tendons have the same strength (per unit cross sectional area)
in most organisms studied. Your bone strength (per unit cross sectional
area) is the same
whether you're an elephant or an elephant shrew. These properties
make sense (perhaps 'collagen is collagen', 'muscle is muscle')
but there are other more complex scale independent properties
that we don't fully understand! Red blood cells (and capillary
diameters) are much the same whatever your body size. Several
other body characteristics vary little with the weight of the
individual.
- physical constraints are things that limit
how much an organism can be modified to cope with changes
in scaling. For example, as Schmidt-Nielsen
has pointed out so well, you and a flea can perform
a standing jump to more or less the same height (The flea can
jump 20cm high, you can raise you centre of gravity about 40cm
jumping from standing. The only reason why you win is because
the flea suffers so much air resistance). But because muscles
can only contract so fast (a physical constraint associated
with the way muscle works) the flea jumps not by simply contracting
its muscles, but by compressing a chunk of highly elastic
resilin, and then explosively releasing this. At such a small
scale, the muscles simply wouldn't contract fast enough to
propel the flea upwards - so the flea has found a trick that overcomes
the physical constraint.
Examples of constraints with scaling implications are:
- Diffusion is hopeless for transporting anything over more
than a tiny distance
- The solubility of oxygen in water is poor
- Nerve conduction speeds are limited by physical factors
(although "higher" organisms have overcome this to a degree
by "re-designing" - developing myelin sheaths)
- Air resistance becomes important as you get smaller
- Don't forget blood viscosity!
- For complex reasons, it appears difficult to "design"
a heart that beats at more than about 1300 beats per minute.
- and so on..
It is intriguing to determine what the physical constraints are
on a complex biological system, but it's even more fascinating observing how
nature manages to sidestep such obstacles!
Allometric Scaling Equations
The magnitude of many body processes changes in a regular fashion as the size of the
organism changes. A surprising number of such processes can be described
in a very simple fashion by:
where M is the body process in question (for example metabolic rate),
x is a measure of the size of the organism, and a and k are constants.
For example, a large number of measurements suggest that the relationship
between metabolic rate (let's call this Pmet) and weight (mb) is:
.. where P is measured in kcal/day and mb is the weight in kilograms.
This equation, first proposed in 1932 by the brilliant animal researcher
Max Kleiber has been amply justified,
and is good for men, mice and elephants! There is a small amount of
uncertainty about the exact exponent (his original equation actually
had an exponent of 0.74) but the value is certainly extremely close to
3/4. There are a whole host of other similar relationships, and many of them
depend on similar "fractions involving a quarter" for example 3/4 and 1/4.
But before we look at these, let's recall that if we take the logarithm
of both sides of an equation, our original equation becomes:
log(M) = log(a) + k * log(x)
You can see that this equation (basically y = c + k*z) is the equation
of a straight line. What does this tell us? Simply, that if we want
to visualise what's going on in one of the above equations, we
can plot log(M) against log(x), and we should get a straight line!
We can also indulge in other mathematical meddling - draw a regression
line through this log-log plot, draw in the 95% confidence intervals,
and the slope of the line will be k, and the point where it meets the
y axis will be log(a) (that is where log(x)=0, in other words where x=1).
 Max Kleiber
Max Kleiber
A list of allometries
In the past fifty years, many people have come up with interesting
allometric equations. Let's have a look at some of these!
(Most data come from Schmidt-Nielsen, and apply to mammals). The lists
look intimidating, but it's worthwhile browsing through them quickly,
for we refer back to them in later discussion. While looking
through, continually ask yourself "Why should this be?".
Little change with weight
- Maximum functional capillary diameter changes little with weight
- Red cell size bears no relationship to weight
- Haematocrit is pretty constant at about 0.45
- Plasma protein concentrations vary little with size or species
- Mean blood pressure is about 100 mmHg more or less independent of size
or species
- Fractional airway dead space (VD/VT) is
pretty constant at about a third
- Body core temperature is weight-independent
- Maximal tensile strength developed in muscle is scale invariant
- Maximum rate of muscle contraction appears scale invariant
- Mean blood velocity has been calculated to be
proportional to mb-0.07
(not much variation)
Changes in (almost) direct proportion to weight
- Heart weight (g) = 5.8 * mb0.98
- Lung weight (g) = 11.3 * mb0.98
- Tidal volume (ml) = 7.69 mb1.04
- Vital capacity (ml) = 56.7 mb1.03
- Lung compliance (ml/cmH2O) = 1.56 mb1.04
- Blood volume (ml) = 65.6 mb1.02
- Muscle mass = 0.40 * mb1.00
- Skeletal mass = 0.0608 * mb1.08
Note how none of the above has an exponent that substantially deviates
from one - as the weight increases, so does the blood volume, the size
of the heart and the size of the lungs. Why should this be, if the
metabolic rate is only increasing with the three-quarter power of the
weight? Note that we don't have an equation for heart stroke volume (do you?)
but it's reasonable to assume that it's directly related to heart
weight.
Scaling to the quarter power
- Heart rate (min-1) = 241 * mb-0.25
- Blood circulation time (seconds) = 17.4 mb0.25
- Respiratory rate (min-1) = 53.5 mb-0.26
Aha! Now we're getting somewhere. If metabolic rate is increasing as
stated, and heart stroke volume (and lung tidal volume) are increasing
in proportion to mass, then something has to give, and thus respiratory
rate and heart rate decrease with increasing size. (You might wish
to ask the question "So why not keep the heart rate the same, and have
a proportionally smaller heart?")
Scaling to the three quarter power
- Pmet (kcal/day) = 73.3 * mb0.75
(Our old friend Kleiber)
- VO2max (ml/s) = 1.94 mb0.79
- Glucose turnover (mg/min) = 5.59 * mb0.75
- In fact, most metabolic parameters vary with the 3/4 power of the
weight. The remarkable implications of this are explored below.
Note that the units of VO2max and Pmet
are different. If we recalculate using the same units, we find that
the VO2max is consistently about ten times the
Pmet.
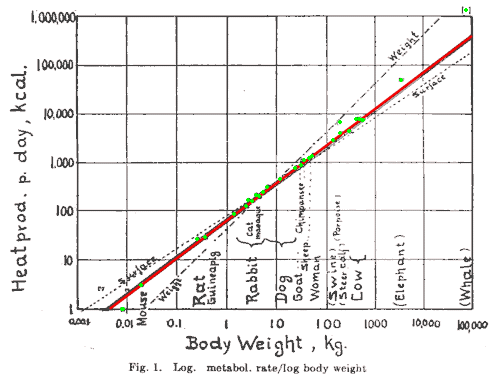
|
A log-log plot of metabolic heat production against body weight, taken
from Kleiber's 1947 article. It is clear that most points (green) closely
fit the red line with a slope of three quarters, rather than the weight line
(slope of 1) or the surface line (slope of two thirds). Note that
although the differences between the lines seem small, this is a log-log
plot so the deviations are actually vast!
Do you wonder why the whale is
where it is?
|
Other allometries
- The metabolic cost of running (ml O2 g-1km-1) =
8.61 * mb0.65
- Mammal brain weight (excluding primates) = 0.01 * mb0.70
- Monkey brain weight = (0.02 to 0.03) * mb0.66
- Ape brain weight = (0.03 to 0.04)* mb0.66
- Brain weight in man = (0.08 to 0.09) * mb0.66
Hmm, okay we just put these in for fun, and to show that not everything
is based on quarter powers. Note that Schmidt-Nielsen gives
a good explanation for the approximately 2/3 exponent in the above
allometric equation for the metabolic cost of running (Others have found
somewhat different exponents).
Using allometric equations
"The above is all very well" you say "but what is the practical use of
all this bumf?" Some of the immediate practical implications are:
- Using 3/4 power scaling
Have you noticed that for some drugs (an example is neuromuscular
blocking agents) per kilogram dosing seems to be wholly appropriate,
while for others, giving a dose based on the body weight is inadequate
at low body weights (for example, aminophyllin and many antibiotics, administered
to children)!
Careful consideration may lead you to the conclusion that the initial
dose of a neuromuscular blocker may depend on blood volume (which scales
with weight) but if the metabolic removal of a drug is important (for
example, aminophyllin) then three-quarter power scaling will apply.
It thus seems reasonable to relate certain drug doses to the 3/4 power
of the weight, and not to either weight or surface area. This isn't
as difficult as it sounds. The cheapest pocket calculator usually has
a square root function and a memory, so if we know the dose for a 70kg person,
let's call it D70 and our patient weighs say 32kg, we
could work out the relevant dose D32
as follows:
This looks formidable but we can easily find the three quarter power
of 32/70 as follows:
- Cube 32/70:
- Clear memory and then work out 32/70
(
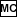 32
32  70
70  0.457 )
0.457 )
- Store the result in memory (
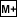 )
)
- Multiply the result by itself, and then by the value retrieved
from memory.
( 0.457  0.457 0.209
0.457 0.209
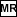 0.0955 )
0.0955 )
- Find the fourth root of the result - press square root twice.
(
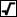 giving the result we want: 0.556)
giving the result we want: 0.556)
You should
not go out and apply this willy-nilly for all drugs, and for
all people. Body composition and behaviour vary remarkably within the
human species, especially at the extremes of size and age. Three
quarter power scaling goes a long way towards explaining why we encounter
statements in the medical literature about the peculiar doses we need to
give to children, and very small or very large adults. If you're dealing with
a metabolic variable (such as a drug dose, a clearance, cardiac output,
or whatever) think Kleiber!
Now that you know this, how about taking the per kilogram doses
you use for children and comparing them with the per kilogram doses
recommended for adults and seeing whether they square up? A really keen
person might like to compare veterinary doses of drugs (for example,
fentanyl) with human doses.
The justification for the above formula is that if
D70 = a*703/4 and D32 = a*323/4
then D32 / D70 = (323/4) / (703/4)
which gives us D32 = D70 * (32/70)3/4.
- Read "the literature" with caution
We have already encountered our dimensionally incorrect friends
Harris and Benedict. Similar circumspection could perhaps be applied to
all reports where for example a metabolic parameter is related to either
weight or to surface area. The likelihood of a major difference is probably
small when dealing with humans (except at the extreme ranges of body mass)
but you should at least feel uncomfortable with information where an
empiric index is used.
For the record, Kleiber reformatted the Harris-Benedict equation (using
their data) in a less dimensionally-challenged way:
- For men: Q = 71.2 * mb3/4
* (1 + 0.004(30-y) + 0.010(S-43.4))
- Women:
Q = 65.8 * mb3/4
* (1 + 0.004(30-y) + 0.010(S-43.4))
Where Q is heat production in kcal/day, w = weight in kilograms,
y = age in years and S, the specific stature, is
height in centimetres / mb1/3
Clearly, it's easier to use the incorrect version, but with modern
calculators and computers, we have little excuse.
- The really good stuff
The real applicability of scaling will probably only become apparent
when we understand what it's all about - why for example as size increases
the heart rate slows, in the place of decreasing cardiac size relative to
weight. Similar slowing may be intimately associated with the lifespan of the organism,
which increases with mass. The lifespan of captive mammals has been found
to approximately fit the allometric equation:
The corresponding equation for birds is
Obviously these are broad rules, with several exceptions. Most animals
seem to survive for about 800 million to a billion heartbeats - humans
are exceptional in this regard, as you can easily calculate. We don't
know what causes ageing, but it's tantalising that the exponent in the
above equations is so close to a quarter. It's not unreasonable to assume
that faster metabolisms cause faster metabolic wear-and-tear on cellular
structures (mitochondria, telomeres and so on), and that this may give
us fundamental insights into ageing.
Quarter-power scaling has even been reported in plants and microbes!
(References: Niklas KJ Am J Bot 1994 81 134 and
Hemmingsen AM Rep Steno Mem Hosp 1950 4 1, ex West 1997).
Explanations Please!
There have been many attempts to explain all, or at least some of the
allometric equations we presented above. If we re-visit our friends
Sarrus and Rameaux, we see that their argument was that as weight
increased so metabolic rate would increase at a lesser rate, depending
on surface area. In other words, they effectively proposed an allometric
equation with an exponent of 2/3 relating metabolic rate and weight.
Although this is not the case, it's not intuitively obvious why they
are wrong.
Some still argue
that the interspecific rule has an exponent of 3/4 but that within
the species the exponent is 2/3. There is little evidence for this,
and the primitive surface area rule also doesn't take into account things
such as fur (which tends to be longer and a more effective insulator
in larger animals, excluding really large tropical animals), and the
thermal conductance of animals, which decreases with increasing size.
( The practical assessment of surface area is in any case a nightmare -
do you include the area of the ears (about 20% of a rabbit's surface area),
the skin area between the legs and the body and so on?)
In some simple circumstances, we have reasonable explantions for
allometric equations. Take the case of pores in the egg of a bird -
as the size of the egg varies, so does the thickness of the egshell,
and total pore area is accurately adjusted to ensure that evaporative
water loss is constant for all eggs (about 15% of weight). Once
we examine more complex cases, our logic tends to break down.
Some attractive explanations for allometric equations must be rejected
out of hand. For example, if we were to predict that the primary determinant
of bone mass was the static load placed on the animal, then the exponent
should be 1.33 and not the 1.08 we
noted above. It is clear that static
loads do not determine bone mass - the important consideration appears
to be stresses on the skeleton during movement! It is often easier to
reject an explanation based on allometric equations, than to accept it
- Karl Popper would be delighted!
"Why three quarters?"
There have been several attempts to explain the 3/4 exponent that
seems to govern metabolism when related to weight. None appears entirely
convincing. Here are a few:
- McMahon (1973)
This argument is based on engineering of the skeleton. He argues
that:
- Animals cannot remain isometric with increasing size,
as loads would increase more than the ability of the skeleton
to withstand such loads;
- All proportional changes in bones should be similar,
as different maneuvers (running, standing etc) impose different
stresses
- The critical length (Lcr) at which a column buckles
is given by:
where d is the diameter, E is Young's elastic modulus, and
rho is the density of the material. This holds for both hollow
and solid cylinders (k differs). In other words, L3
is proportional to d2
- If the weight of a limb ml is proportional to the
body weight mb, then
mb is proportional to L * d2
(as the limb weight is proportional to length (L) times diameter (d) squared)
- But by our elastic criteria, L3 is proportional to
d2, so mass is proportional to L4, which
is another way of saying that L is proportional to mb1/4.
Similarly d is proportional to mb3/8.
- Because work is force * distance, the power output of a muscle
is given by
Pmax = maximal tensile stress * area * change in length / time
Now the maximal tensile stress in a muscle is scale-invariant,
as is change in length per unit time. Thus muscle power depends
only on cross sectional area. If this area is
proportional to d2 (a reasonable assumption) then
which conveniently works out to mb3/4
I would like to see a good critique of this theory, which certainly
seems to work in cattle, but the general applicability has been questioned.
- West et al (1997)
These authors used an extremely complex argument to achieve the
magical three quarters. They use an entirely different tack from that
of McMahon - they look at the energy cost of transporting essential
materials to where they are needed.
Their basic assumptions are that minimal energy is dissipated
(not unreasonable) and that terminal capillaries are size-invariant
(we know this). Transport is through a space-filling fractal network
of branching tubes and the model has fairly general applicability.
They argue that:
- The metabolic rate is proportional to the flow (Q) through the
system, and that at the kth level of branching,
Q = Nk.pi.rk2uk, where
Nk is the number of branches at the kth level, and
uk is the mean flow velocity (averaged over area and time).
Specifically, for capillaries, Q = Nc.pi.rc2uc.
- They assume that capillary radius, length and mean flow (abbreviated
rc, lc and uc) are
size invariant. If the allometric equation governing the relationship
between Q and body mass mb is Q = k.mba,
it follows that Nc scales in proportion to mba.
In other words, the number of capillaries scales in proportion to the
metabolic rate.
- They prove that for minimal energy dissipation, the branching of
the network must follow the rules for a self-similar fractal. In this
circumstance, the number of branches increases in geometric proportion
as their size decreases. If each tube at branching level k branches
into nk smaller ones, then for a self-similar fractal,
Nk = nk ie as k increases so the number of
branches increases in geometric proportion. Thus, as the mass increases,
so the number of capillaries increases with the logarithm of the mass.
- They use the above to prove that a, the exponent of mb
in the allometric scaling equation, is equal to ln(n)/ln(gamma.beta2),
where gamma and beta are scale factors that characterise the branching
of the system (In general for such systems betak = rk+1/rk and
gammak=lk+1/lk, but in these circumstances,
the values are constant at all levels, and simply referred to as beta
and gamma).
- Further complex argument (assuming that branching is area-preserving)
gives the magic number of a = 3/4. Woops, in mammalian systems, the
branching isn't area-preserving, but all is not lost, as yet more complex
arguments again give us the magic 3/4 by invoking pulsatile flow as a
saviour.
I have a lot of trouble with the above argument.
-
The main tenor of
the argument seems to be that the three-quarter power is a necessity
imposed by the fractal nature of the system, held in place by the
necessity to minimise the energy used in distributing nutrients to
the tissues. They overlook the fact that the energy required by the
pumping action of the heart is a relatively small proportion of total
energy expenditure. (Note that as Schmidt-Nielsen details, the oxygen
cost of supplying 1 ml of oxygen to the tissues is
probably size invariant).
- Even more worrying about the above argument is the implied assertion
that the nature of the "fuel delivery" system constrains the organism
to have a higher metabolic rate than we would expect from the simple
necessity to balance energy production and loss (our old friends
Sarrus and Rameaux). Guyton's argument that the peripheral tissues
regulate cardiac output by their demands (rather than central control)
seems to stand in the way of this sort of reasoning, as does the evidence
that vessel formation is dictated by the needs of the tissues, rather
than the other way around.
- The argument overlooks the relationship between an animal and
its environment. The major constraint on energy utilisation by an animal
is surely whether it is using that energy optimally in relation to its
environment. Most animals are not standing around tamely in cages
having their metabolisms measured by physiologists - they are interacting
with their environment - hunting or escaping, feeding or migrating,
shivering or panting, and so on. Any argument that ignores this is
surely incomplete by its very nature!
- Banavar & colleagues
This argument is also complex - it too is based on the concept of
a branching network supplying nutrients, as is that of West. The
argument here is much more general, with fewer constraints. They
too derive a three quarter exponent by:
- Proving that for the most efficient network, blood volume C
scales with LD+1 where L is a parameter of linear size, and D the
number of dimensions of the object (3, in our system).
- Asserting that blood volume scales with mass;
- Proving that metabolic rate (B) scales with LD
- And thus, as mass is proportional to LD+1 and
L is proportional to (metabolic rate)1/D we get our magic
result that metabolic rate is proportional to massD/D+1
Hmm. I'm afraid that despite its apparent greater generality,
this argument too is susceptible to the problems we found with
West's one.
- Witting
Witting does use an environmental argument, and he too arrives
at the magical 3/4 exponent, but only for terrestrial animals
that forage in two dimensions. For pelagic ones (that forage in three
dimensions) the exponents should be different. Hmm - so far the
evidence seems to be in his favour, but we watch eagerly for more
hard data. You may wish to check out his
 web-site where some downloads are available.
web-site where some downloads are available.
- Nobody
Nobody seems to have fully considered that an important constraint on
animal survival is it's peak capacity for work (rather than its so-called
"basal" energy output), although McMahon has touched on this.
If you are either a predator or prey, you need to
be able to run bloody fast at times, or you will starve or be eaten!
Interestingly enough, the principles of symmorphosis here seem to apply
to the whole organism. Consider how top-class athletes are continually
damaging themselves - ruptured tendons, torn muscles, broken bones and
so on seem to be the inevitable concomitants of pushing oneself to the
limit - as if the whole organism were designed around these maximum
tolerances. (In contrast, there has been much argument that the
respiratory system does not fit in with the principle of
symmorphosis!)
As we have noted
above the maximum oxygen consumption of an
organism appears to usually be about ten times its basal consumption
(some have termed this "factorial aerobic scope"). The question then
arises "If the maximum oxygen consumption of an organism is x, does
this maximum necessitate some corresponding minimum oxygen
consumption (perhaps x/10) that we have graced with the term 'basal
oxygen consumption'?" In other words, have we got things the wrong
way around? We should carefully consider those organisms with a factorial
scope of over 10 (for example, highly trained athletes and dogs, who
have a factorial scope of about twenty) and
see how they fit into the picture! Have you noticed how smaller dogs
seem to live longer than larger ones, apparently defying Kleiber's rules?
References
- Kleiber M. Physiological Reviews 1947 27 511-541.
A brilliant review. Read it!
- Taylor and Weibel. Respiratory Physiology 1981 44 1-10.
- Schmidt-Nielsen K, Scaling. Why is animal size so important?
Cambridge University Press, 1984. ISBN 0 521 31987 0,
ISBN 0 521 26657 2. Get the book. A fascinating read! Much of
the above is based on this book.
- McMahon TA Science 1973 179 1201-4
- West GB, Brown JH and Enquist BJ. Science 1997 276 122-6.
- Banavar JR, Maritan A and Rinaldo A. Nature 1999 299 130-2
and Supplementary information at
nature.com
- Heusner. Respiratory Physiology 1982 48 1-12.
Arguments in favour of a 2/3 exponent within a species.
Web References
Take a look at: